Väitöskirja Petersen Anna.
Oxidative stress and proteolysis in the lens- effects of
anti-inflammatory drugs. ISBN
91-628-6746-6 Väitöstilaisuus
10.3.2006 Sahlgrenska Akademi Göteborgs universitet.
Silmän linssin oksidatiivinen stressi ja proteolyysi- Anti-inflammatoristen lääkkeitten vaikutus.
Abstraktin suomennosta
TAUSTA: Oksidatiivisella
stressillä tiedetään olevan merkitystä iän mukana kehittyvän
kaihin synnyssä.
TARKOITUS:Tämän
väitöskirjatyön tarkoituksena on ollut selvittää proteolyysin
redox-säätelyä ja anti-inflammatoristen lääkeaineitten
oksidatiivisia ja antioksidatiivisia vaikutuksia ihmisen silmän
linssin epiteelisoluissa (HLEC) ja hiiren intaktisssa linssissä.
MENETELMÄ: Tutkimuksisssa
on nesteessä ollut fysiologiset olosuhteet, sokerin pitoisuus on
vastannut 5,2 mmol. Sokeri ei ole ollut muuttujana. Ihmisen linssin
epiteelisolut ja intaktit hiiren linssit altistettiin
NSAID-lääkkeille kuten indometasiinille, diklofenakille,
celekoksibille ja asetyylisalisyylihapolle, samoin glukokortikoidille
dexametasoni. HLEC-soluihin kohdistettiin oksidatiivinen stressi
(H2O2). Apoptoosia tutkittiin solutumista Hoechstin
värjäysmenetelmällä ja kaspaasi-3 aktiviteettia mittaamalla.
Soluihin järjestettiin superoksidianioinin (O2-)
vaihtelevaa tuottumista (dihydroetiniumilla), peroksidien
vaihtelevaa tuottumista
(diklorofluoresiinidiasetaatilla) ja glutationin (GSH) vaihteluja (
monoklorobimanilla). Mitokondrioitten depolarisaatiota mitattiin
jännitteelle herkällä värillä JC-1. Morfologiaa tutkittiin
transmissio-elektronimikroskopialla (TEM). Kalpaiinin, proteosomin ja
happamien lysosomaalisten proteaasien proteolyyttinen aktiviteetti
mitattiin käyttämällä fluorogeenisia substraatteja.
TULOKSET: Matalapitoinen
NSAID ja ASA suojasivat vetyperoksidin (H2O2)
indusoimalta apoptoosilta HLEC-soluja, kun
taas korkeat pitoisuudet olivat toksisia. Matalapitoisina NSAID ja
ASA vähensivät superoksidianionin (O2-)
ja peroksidin tuottumista
sekä GSH:n vähenemistä oksidatiivisen stressin alaisissa
HLEC-soluissa. Mutta ei voitu havaita mitään NSAID:n ja ASA:n
antamaa suojaa vetyperoksidin (H2O2)
indusoimaa mitokondriamembraanin
depolarisaatiota vastaan.
Deksametasoni lisäsi huomattavasti apoptoosia, mutta sillä ei
ollut mitään vaikutusta superoksidin (O2-)
tuottoon, GSH:n pitoisuuteen tai mitokondrian membraanipotentiaaliin.
Vain korkein deksametasonipitoisuus (100 mikromoolia) osoitti
kohonnutta peroksidin tuottoa. TEM-menetelmällä havaittiin
deksametasonille altistettujen HLEC-solujen järjestyminen moniin
kerroksiin, vakuolien muodostuminen ja elektronitiheitten
multivesikulaaristen kappaleitten ilmestyminen näihin
linssiepiteelisoluihin.
Kehitettiin uusi metodi mitata hiiren intaktin
linssin proteolyysi. Proteolyyttistä aktiviteettia stimuloitiin
solun sisäistä kalsiumpitoisuutta kohottamalla. ( ionomysiinin
lisäyksellä). Proteosomin inhibiittori lactacystiini
vähensi huomattavasti proteolyysiä. Sen sijaan kalpaiinin
inhibiittorit ja lysosomaalisten entsyymien inhibiittorit eivät
vähentäneet proteolyysiä. Kaikki proteosomin kolme pääpeptidaasia
HLEC-lysaatissa vaikuttuivat, kun niitä inkuboitiin joko
redusoidulla (GSH) tai oksidoidulla (GSSG) glutationilla;
Kymotrypsiinin kaltaiset ja peptidyyliglutamyyli-peptidaasin
kaltaiset aktiviteetit antoivat samankaltaisen vasteen kun lisättiin
GSH:ta. Mutta trypsiininkaltainen aktiviteetti inhiboitui, kun GSH
lisättiin. Mutta kun lisättiin GSSG, kaikkien kolmen peptidaasin
aktiviteetti inhiboitui. Mutta jos samanaikaisesti inkuboitiin
redusoivan aineen ditiotreitolin
kera, kumoutui
GSSG:n estovaikutus.
Vetyperoksidin (H2O2)
aiheuttama oksidatiivinen stressi vähensi huomattavasti
HLEC-soluviljelmissä proteosomin pääasiallista
peptidaasiaktiviteettia. Kun intakteja hiiren linssisoluja
inkuboitiin vetyperoksidin vaikutuksen alaisina, havaittiin
proteosomissa taipumus kymotrypsiiniaktiviteetin vähenemiseen ja
alentuneisiin GSH-pitoisuuksiin.
JOHTOPÄÄTÖKSENÄ
on,
että NSAID ja ASA (VAIN matalin pitoisuuksin) suojaavat apoptoosilta
oksidatiivisen stressin alaisia ihmisen linssin epiteelisoluja
antioksidatiivisin aktioin. Steroideille altistuneet ihmisen linssin
epiteelisolut osoittivat soluviljelmissä solujen kertymistä
kerroksiin ja apoptoosin lisääntymistä, mikä viittaa
häiriintyneeseen differentiaatioon, häiriintyneeseen
proliferaatioon ja häiriintyneeseen migraatioon, mitkä ovat
mahdollinen mekanismi steroidien aiheuttaman kataraktan
kehittymisessä. Solun sisäinen redoxstatus säätelee tiukasti
linssin epiteelisolujen proteolyysin.
(Tekstiosasta:
Solujen GSH/GSSG-suhde on tärkeä proteosomin
säädössä. 20S proteosomi on osana ubikitiini-proteosomitietä
(UPP), jota pidetään tärkeänä oksidoituneitten tai muuten
vaurioituneitten, iän mukana kehkeytyvien proteiinien
poistamisessa. Oksidoituminen on tavallinen ikääntymisilmiö,
samoin vetyperoksidin lisääntyvä osuus kaihiintuvassa silmässä-
jolle C- ja E-vitamiini taas ovat vastavaikuttavia ja suojaavia
antioksidantteja). Silmän linssillä on useita suojajärjestelmiä
oksidatiivista stressiä vastaan , eräs tärkeimmistä on
glutationisysteemi, GSH. Korkea GSH-pitoisuus voi ylläpitää
linssiproteiineissa reduktiotilaa ja samalla essentiellisti suojata
sisä- ja ulkosyntyisiä vapaita happiradikaaleja vastaan).
Suomennos 10.3. 2006 lb
Suomennos 10.3. 2006 lb

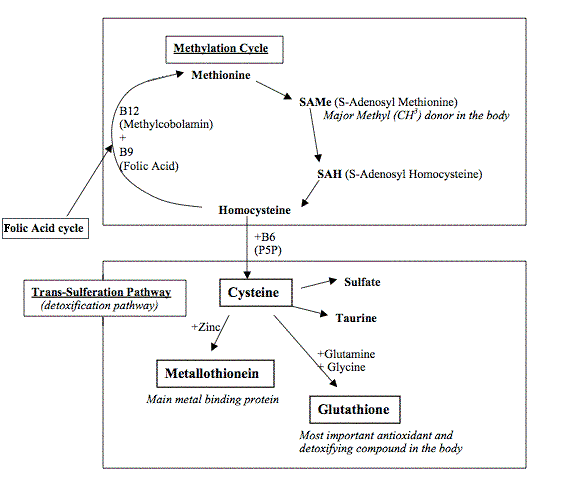
 Kuten kartasta huomaa, täydellisen kartan kärhenä on tauriinin normaali muodostus.Tauriini on tavallaan merkitsijä rikkiaineenvaihdunnasta.
Kuten kartasta huomaa, täydellisen kartan kärhenä on tauriinin normaali muodostus.Tauriini on tavallaan merkitsijä rikkiaineenvaihdunnasta.